分裂組織パラメータ
(Meristem parameters)
分裂組織(Meristem)セクションのパラメータは、分裂組織(meristems)がどんなふうに働くのか、植物がどんなふうに分枝し、発達していくのかを決めるものです。
腋生分裂組織と葉序
(Axillary meristem and leaf arrangement)
植物はしばしば、各節間につく葉の枚数で分類されます。1枚のものを互生(alternate)、2枚のものを対生(opposite)、3枚以上のものを輪生(whorled)といいます。PlantStudio は、互生と対生をシミュレートします。ほとんどの植物が、互生か対生のどちらかです。互生と対生のどちらを選ぶかによって、分枝や花房形成に利用可能な腋生分裂組織の数がきまります。腋生分裂組織は、葉と茎(軸)とに挟まれる部分に形成されるからです。そのため、対生の葉序を持つ植物は、互生の葉序を持つ植物の2倍の腋生分裂組織(わき芽)を持つことになります。
分裂組織の3Dオブジェクト、大きさ、回転、色
(Meristem 3D object, scale, rotations, colors)
これらのパラメータは、分裂組織(芽)が植物にどのように現れるかを決めるものです。通常は、分裂組織は表示したくないでしょうから、[分裂組織パラメータ:分列組織の最大時のサイズ(Meristems: Meristem 3D object scale at full size)]パラメータは0に設定されています。しかし、分裂組織がどのように働いているのかをみたいなら、あるいは、ある種の植物のように小さな芽を付けたいなら、この大きさの値を増加させます。
分枝指数、頂芽優勢の強さ、二次分枝
(Branching index, Apical dominance strength, Secondary branching)
PlantStudio での分枝は次のように行なわれます:まだ生殖生長を行なっていない腋芽は、毎日、新しい枝を作り始めるかどうかを考えます。この決定にあたっては、次の3つの事柄が斟酌されます。
- まず第一に、植物全体にその効力が及んでいる分枝指数(branching index)を考慮します。分枝指数は、0から100の間に設定されています。この値は、分裂組織(芽)が枝になる可能性の度合を示しています。各分裂組織(芽)では、0から100の数値がランダムに作り出され、この値が分枝指数よりも小さいと、その芽は新しい枝を作り始めます。
- 次に、その分裂組織(腋芽)は、頂端分裂組織(頂芽)との距離に応じて、近ければ近いほど、そこで用いる分枝指数を低くします。植物が自身の姿をコントロールする方法の一つに頂芽優勢(apical dominance)があります。頂芽優勢においては、その植物の茎の頂端にある頂芽が、側芽の分枝を抑える物質を送ることによってそれら側芽を支配します(dominate)。(園芸の本で、枝を育てたい植物では「上部を刈り込む(top)」ように書いてあるのはこの理由によります。この作業で頂芽を取り除いているのです。)PlantStudio は、頂芽に近いところにある芽が分枝する確率を低めることで頂芽優勢をシミュレートします。[分裂組織パラメータ:長が優勢の強さ(節間の長さで指定)(Meristems: Apical dominance strength (as node distance))]パラメータを設定することで頂芽優勢が始まる地点を制御できます。例えば、節間が5つの植物があったとして、頂芽優勢の強さ(apical doninace strength)を 3.0 に設定すると、下から2つの節間は普通に分枝するけれど、(頂芽に近接する)残り3つの節間は、分枝しずらくなります。
- 最後に、二次分枝をオフに設定していて、その枝が植物の主茎でない場合、その枝にある分裂組織は分枝できません。
通常、分枝指数は30%より小さい値に設定しておくのがよいでしょう。それより高い値に設定すると、枝がたくさんになりすぎて描画に時間がかかるようになるからです。分枝する能力のある腋芽の数を変更することで分枝の量を制御することもできます。葉序の互生(alternate leaf arrangemant)を選び、また、新規の節間と葉を作るのに必要な最小日数を増やすことで、腋芽の数を少なくすることができます。
仮軸分枝
(Sympodial branching)
植物の多くは、単軸分枝です。単軸分枝では、頂芽が主茎を伸ばし、側芽は新しい枝を作ります。仮軸分枝においては、頂芽は節間を一つ作った後、死んでしまい、その頂芽が作り出した側芽の一つが次の主茎を作っていきます。このタイプの分枝は、トマトでみることができます。このパラメータをオンにすると、仮軸分枝になります。次のことに注意してください: 仮軸分枝を選択した場合、腋生分裂組織と葉序のパラメータで対生(opposite)を選択していないと、分枝指数を変更しても何の効果もありません。なぜなら、(互生だと)腋芽のすべてが主茎の生成に使われてしまうからです。
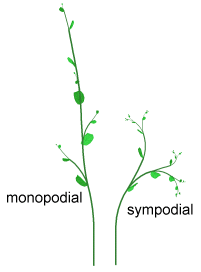
[訳注:
左図:単軸分枝、右図:仮軸分枝]
分枝角度
(Branching angle)
分枝角度は、新規の枝が、それまで作ってきた枝(茎)との間に作る角度です。PlantStduio は重力モデルを持たないので、このパラメータ値をあまり大きく設定すると、ありえない角度で枝ができてしまいます。ですから、分枝をあまりしないとか、二次分枝を許可しない、というような場合以外では、この値を90度以下にしておくとよいでしょう。
開花信号を受け取った時に、各分裂組織が生殖成長を始める確率
(Probability each meristem will become reproductive when gets flowering signal)
このパラメータは、決定確率(determinate probability)とも呼ばれます。「決定した植物(determinate plants)」は、生殖生長を始るようになり、すべてのことが生殖生長へと注がれ、栄養生長へと再び戻ることは決してありません。一方、「決定していない植物(indeterminate plants)」は、リソースの多くは生殖へと振り向けられているけれど、まだ栄養生長をします。トマトの品種でこの違いを見ることができます:低木性のトマトは、「決定した植物」である場合が多く、生長してから花をつけ、滅びていきます。一方、つる性のトマトは通常、「決定していない植物」で、生長し、次に花をつけまた生長し、また花をつけさらに生長し、環境さえ整えば長いこと生き続けます。PlantStudio は、決定確率を使ってこの違いをシミュレートします。決定確率は、その植物が生殖成長相に入る時に、ある分裂組織が「決定する(determinate)」確率です。この決定確率が1ならば、その植物はこれ以上の栄養生長部分(葉や茎など)を作りません。決定確率0ならば、その植物は決して花をつけません。0.5ならば約半分の分裂組織が生殖成長へとスイッチを切り替え、残りの半分は栄養成長を続けます。